Cyklická kontinuita živých bytostí nachádza väzby medzi postupnými generáciami vo fenoméne reprodukcie.
Reprodukcia sa vykonáva na rôznych úrovniach evolučného rozsahu, v rôznych dôsledkoch rastlinnej a živočíšnej ríše, v rôznych živých druhoch, s takou rôznorodosťou mechanizmov, ktoré samotné odôvodňujú celé pojednanie.
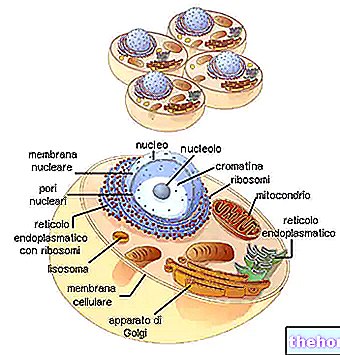
Prvá klasifikácia reprodukčných javov musí odlišovať jednobunkové od mnohobunkových organizmov, pretože iba v prvom z nich sa delenie buniek zhoduje s reprodukciou.
V mnohobunkových môže byť reprodukcia agamická alebo sexuálna (alebo keramická).
Relatívne menej častá agamická reprodukcia je založená na mechanizme mitózy, takže variabilita druhu je skôr zverená opakovanému výskytu mutácií.
Existujú aj rôzne mechanizmy, ako je strobilizácia, sporulácia atď., Zatiaľ čo v rastlinách nachádzame regeneračné formy dobre známe v poľnohospodárstve (odrezky, vrstvenie atď.).
Najrozšírenejším reprodukčným mechanizmom vo vyšších formách je však sexuálny, ktorý zodpovedá vzhľadu meiózy, tvorbe gamét a ich fúzii v zygote (oplodnenie).
U primitívnych druhov nie sú gaméty morfologicky diferencované: v tomto prípade hovoríme o izogametii. Vždy však existujú dve série gamét, identifikované symbolmi (+) a (-) a oplodnenie môže nastať iba zo „stretnutia medzi gamétami opačného znamienka: existuje teda biologický rozdiel, ktorý ešte nie je vyjadrený morfologicky.
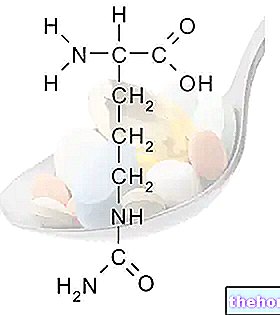
Ako evolučná škála pokračuje, objavuje sa morfologická a funkčná diferenciácia, v ktorej je typ ženských gamét spravidla obdarený bohatým rezervným materiálom (deutoplazma alebo teľa, ktoré zabezpečia vývoj embrya, až kým nebude metabolicky nezávislý) a typ mužské gaméty vybavené pohyblivosťou dosiahnuť ženské. Gamety sú vždy haploidné a sú výsledkom meiózy. Ich zlúčením vzniká diploidná zygota.
Medzi meiózou a zygotou môže uplynúť séria generácií haploidných buniek, rovnako ako medzi zygotou a meiózou môže uplynúť séria generácií diploidných buniek s rôznymi druhmi generatívnych cyklov vyjadrených striedaním generácií.
Druh diplonte (s diploidným organizmom) je charakterizovaný gametickou meiózou: meióza priamo produkuje gaméty, ktoré zlúčením okamžite rekonštituujú diploidný stav. Toto je rozšírený prípad Metazoanov, vrátane človeka.
Gametogenéza
Keď vezmeme do úvahy reprodukciu s gametickou meiózou, ako sa vyskytuje u človeka, pokúsme sa objasniť, ako meióza zapadá do gametogenézy (tvorba gamét).
V embryonálnom vývoji existuje u mužskej a ženskej gametogenézy (nazývanej spermatogenéza a oogenéza) včasná diferenciácia medzi bunkami určenými na vytvorenie tela (somatická línia) a bunkami určenými na produkciu gamét (zárodočná línia). Počiatočné bunky zárodočnej línie sa nazývajú protogóny. S diferenciáciou gonád v mužskom alebo ženskom zmysle dochádza k diferenciácii zárodočných buniek na spermatogóniu a ovogoniu.
Pri pohľade na spermatogenézu vidíme, že v spermatogónii existuje séria bunkových generácií, ktorá pokračuje po celý život. Len časť takto kontinuálne produkovanej spermatogónie sa líši od normálneho mitotického cyklu a namiesto toho začína meiotická.
Zárodočná bunka, v ktorej začne meióza (zdvojenie a potom prvé delenie), sa nazýva spermatocyt prvého rádu; jeho delením vzniknú dva spermatocyty druhého rádu, z ktorých pri druhom delení vzniknú celkom štyri spermatidy.
Redukciu chromozomálnej súpravy môžeme absolvovať zo 4n spermatocytov prvého poriadku (po zdvojení existujú štyri chromatily pre každý pár homológov) na 2n spermatocytov druhého rádu a na n spermatidov, ako už bolo vidieť štúdiom meióza, ktorú teda uzatvárame. Spermie sú teda už haploidné, ale ešte nie sú zrelými gamétami. Zo štruktúry bunky haploidného typu funkčné dozrievanie (nazývané spermiohistogenéza) transformuje spermatidy na spermie, teda zrelé samčie pohlavné bunky.
V ženskej gametogenéze (alebo oogenéze) existuje niekoľko rozdielov. Po prvé, počet gamét, ktoré je potrebné pripraviť, je oveľa menší. Odhaduje sa, že v gonádach samice ľudského druhu je pripravených asi 5 X 105 ovogónov; z nich len asi 400 má záujem o dozrievanie folikulov a následnú skleslosť, v cykle, ktorý zvyčajne postihne iba jeden folikul mesačne počas plodného obdobia asi 35 rokov.
Rozdielny počet gamét pripravených pre tieto dve pohlavia zodpovedá už uvedenému rozdielu vo funkcii a správaní: spermie sú malé, mobilné a početné v súvislosti s potrebou hľadať vajíčko a nízkou pravdepodobnosťou jeho nájdenia; vajíčka sú veľké, inertné a málo vo vzťahu k funkcii záruky zárodku rezervného materiálu a ochrany, ktorú mu poskytuje vnútorné oplodnenie (prirodzene, najmä pri vonkajšom oplodnení musia byť vajíčka tiež početnejšie).
Potreba poskytnúť gaméty s rezervným materiálom zodpovedá prítomnosti „oogenézy“ fázy zastavenia meiózy, počas ktorej sú chromozómy čiastočne despiralizované. Potom pozorujeme takzvané „perové chromozómy“, v ktorých extroverzií identifikuje znaky, v ktorých sú derepresívne gény zodpovedné za syntézu deutoplazmy.
Menší počet požadovaný pre ženské gaméty tiež zodpovedá skutočnosti, že zo štyroch haploidných buniek produkovaných meiózou iba jeden dostane celý rezervný materiál a stane sa gamétou, zatiaľ čo ostatné tri (polocyty alebo polárne telá), ktoré obsahujú iba chromozomálny materiál, nemôže viesť k zygotom a embryám a je predurčený na regresiu.
Hnojenie
Hnojenie, tj stretnutie medzi mužskou a ženskou gamétou, sa môže uskutočňovať veľmi rôznymi spôsobmi.V živočíšnej ríši pozorujeme prechod od vonkajšieho oplodnenia (gaméty vystavené akémukoľvek environmentálnemu riziku, a preto nevyhnutne veľmi početné u oboch pohlaví) k vnútornému oplodneniu, s ktorým je rodičovská starostlivosť ďalej spojená až k metabolickému vzťahu cicavcov medzi matkou a plodom .
Hnojenie, akonáhle dôjde k stretu medzi gamétami opačného pohlavia, musí prebehnúť tak, aby boli zaručené dve podmienky: špecifickosť a jedinečnosť. To znamená, že musí byť zaistené, aby spermie boli rovnakého druhu ako vajíčko a aby, akonáhle vstúpi prvé, nevstúpia žiadne ďalšie.
Špecifickosť je zaistená biochemickými charakteristikami akrozómu a povrchu vajíčka. V skutočnosti sa hovorí o reakciách medzi «fertilisínmi» a «antifertilisínmi» so špecifickosťou porovnateľnou so špecifickosťou stretnutia medzi enzýmami a substrátom.
Jedinečnosť oplodnenia je zaručená úpravou povrchovej štruktúry vajíčka («kortikálna reakcia»), ktorá začína hneď v okamihu prvej špecifickej reakcie na fertilisín / antifertilisín; po tejto reakcii sa membrána vajíčka zmení, takže akékoľvek iné spermie, ktoré sa k nej dostanú, už nie sú schopné iniciovať špecifickú reakciu oplodnenia.
Po oplodnení zostáva chvost spermotozoanu mimo vajíčka, pričom do neho preniká chromozomálny materiál. Toto, nazývané „mužský pronukleus“, sa spája s „ženským pronukleom“ vajíčka, čím vzniká diploidné jadro zygoty.