Ak chcete hovoriť o dvadsiatich aminokyselinách, ktoré tvoria proteínové štruktúry, a o modifikovaných, bolo by potrebné popísať najmenej dvanásť špecializovaných metabolických ciest.
Prečo však bunky používajú toľko metabolických ciest, ktoré vyžadujú energiu (napríklad na regeneráciu katalytických miest enzýmov), z ktorých každá má enzymatické dedičstvo, na katabolizáciu aminokyselín? Takmer zo všetkých aminokyselín je možné získať špecializovanými cestami metabolity, ktoré sa v malej časti používajú na výrobu energie (napríklad prostredníctvom glukoneogenézy a dráhy ketolátok), ale ktoré predovšetkým vedú k tvorbe komplexných molekuly s vysokým počtom atómov uhlíka (napríklad z fenylalanínu a tyrozínu sa v nadobličkách špecializovaných na tento účel produkujú hormóny); ak by na jednej strane bolo jednoduché vyrábať energiu z aminokyselín, na strane druhej by bolo komplikované stavať komplexné molekuly vychádzajúce z malých molekúl: katabolizmus aminokyselín im umožňuje využiť svoju kostru na získanie väčších druhov.
U zdravého jedinca sa denne odbúravajú dva alebo tri hektogramy aminokyselín: 60-100 g z nich pochádza z bielkovín prijatých v potrave, ale viac ako 2 hektogramy sa získajú z normálneho obratu bielkovín, ktoré sú neoddeliteľnou súčasťou organizmu ( aminokyseliny týchto bielkovín, ktoré sú poškodené redoxnými procesmi, sú nahradené inými a katabolizované).
Aminokyseliny poskytujú energetický prínos z hľadiska ATP: po odstránení a-aminoskupiny môže zostávajúca uhlíkatá kostra aminokyselín po vhodných transformáciách vstúpiť do Krebsovho cyklu. Okrem toho, keď chýbajú zásoby živín a množstvo glukózy klesá, aktivuje sa glukoneogenéza: glukoneogenetickými aminokyselinami sa rozumejú tie, ktoré je možné po vhodných úpravách zaviesť do glukoneogenézy; glukoneogenetické aminokyseliny sú tie, ktoré je možné premeniť na pyruvát alebo vo fumaráte (fumarát je možné previesť na malát, ktorý opúšťa mitochondriu a v cytoplazme sa transformuje na oxaloacetát, z ktorého je možné získať fosfoenolpyruvát). octan octový.
Ten práve popísaný je veľmi dôležitým aspektom, pretože aminokyseliny môžu napraviť nedostatok cukru v prípade bezprostredného pôstu; ak pôst pretrváva, po dvoch dňoch zasiahne metabolizmus lipidov (pretože na štruktúry bielkovín nemožno príliš útočiť) práve v tejto fáze, pretože glukoneogenéza je veľmi obmedzená, dochádza k premene mastných kyselín na acetylkoenzým A a ketolátky. Od ďalšieho pôstu sa mozog tiež prispôsobuje používaniu ketolátok.
K prenosu a-aminoskupiny z aminokyselín dochádza transaminačnou reakciou; enzýmy, ktoré katalyzujú túto reakciu, v skutočnosti hovoria, transaminázy (alebo aminotransferáza). Tieto enzýmy používajú enzymatický kofaktor nazývaný pyridoxalfosfát, ktorý zasahuje do svojej aldehydovej skupiny. Pyridoxalfosfát je produktom fosforylácie pyridoxínu, čo je vitamín (B6), ktorý sa nachádza hlavne v zelenine.
Transaminázy majú nasledujúce vlastnosti:
Vysoká špecificita pre pár ketoglutarát-glutamát α;
Pomenované sú podľa druhého páru.
Transaminázové enzýmy vždy zahrnujú pár a ketoglutarát-glutamát a rozlišujú sa podľa druhého zapojeného páru.
Príklady:
L "aspartát transamináza tj GOT (Glutamate-Ossal acetát transamináza): enzým prenáša a-aminoskupinu z aspartátu na a-ketoglutarát, pričom sa získava oxaloacetát a glutamát.
L "alanín transamináza tj. GTP (glutamát-pyruvát transamináza): enzým prenáša a-aminoskupinu z „alanínu“ na a-ketoglutarát, pričom sa získava pyruvát a glutamát.
Rôzne transaminázy používajú a-ketoglurát ako akceptor aminoskupiny aminokyselín a prevádzajú ho na glutamát; pričom vzniknuté aminokyseliny sa používajú v dráhe ketolátok.
K tomuto druhu reakcie môže dôjsť v oboch smeroch, pretože sa lámu a vytvárajú väzby s rovnakým energetickým obsahom.
Transaminázy sú v cytoplazme aj v mitochondriách (väčšinou sú aktívne v cytoplazme) a líšia sa svojim izoelektrickým bodom.
Transaminázy sú tiež schopné dekarboxylovať aminokyseliny.
Bude musieť existovať spôsob, ako previesť glutamát späť na α-ketoglutarát: to sa deje deamináciou.
Tam glutamát dehydrogenáza je to enzým schopný transformovať glutamát na α-ketoglutarát, a teda prevádzať aminoskupiny aminokyselín nachádzajúcich sa vo forme glutamátu na amoniak. Odohráva sa redoxný proces, ktorý prechádza medziproduktom a-amino glutarátu: amoniak a a-ketoglutarát sa uvoľňujú a vracajú sa do obehu.
Potom likvidácia aminoskupín aminokyselín prechádza transaminázami (ktoré sa líšia podľa substrátu) a glutamátdehydrogenázou, ktorá určuje tvorbu amoniaku.
Existujú dva typy glutamátdehydrogenázy: cytoplazmatické a mitochondriálne; kofaktor, ktorý je tiež kozubstrátom tohto enzýmu, je NAD (P) +: glutamát dehydrogenáza používa ako akceptor redukčnej sily buď NAD + alebo NADP +. Cytoplazmatická forma uprednostňuje, aj keď nie výlučne, NADP +, zatiaľ čo mitochondriálna forma uprednostňuje NAD +. Mitochondriálna forma má za cieľ likvidáciu aminoskupín: vedie k tvorbe amoniaku (ktorý je substrátom pre špecializovaný enzým v mitochondriách) a NADH (ktorý je odoslaný do dýchacieho reťazca). Cytoplazmatická forma funguje opačným smerom, to znamená, že používa amoniak a a-ketoglutarát na získanie glutamátu (ktorý má biosyntetické miesto určenia): táto reakcia je reduktívnou biosyntézou a použitým kofaktorom je NADPH.
Glutamátdehydrogenáza funguje, keď je potrebné zbaviť sa aminoskupín aminokyselín, ako je amoniak (močom), alebo keď sú kostry aminokyselín potrebné na výrobu energie: tento enzým bude mať preto ako negatívne modulátory systémy, ktoré sú indikáciou s dobrou energetickou dostupnosťou (ATP, GTP a NAD (P) H) a ako pozitívne modulátory systémy, ktoré naznačujú potrebu energie (AMP, ADP, GDP, NAD (P) +, aminokyseliny a hormóny štítnej žľazy).
Aminokyseliny (hlavne leucín) sú pozitívnymi modulátormi glutamátdehydrogenázy: ak sú v cytoplazme prítomné aminokyseliny, môžu sa použiť na syntézu bielkovín alebo sa musia zlikvidovať, pretože sa nedajú akumulovať (to vysvetľuje, prečo sú aminokyseliny pozitívnymi modulátormi ).
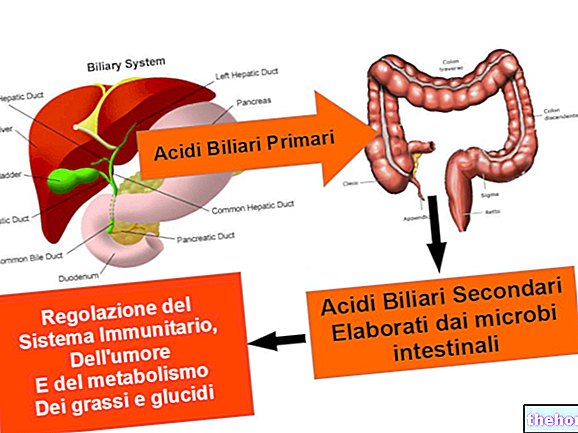
Likvidácia amoniaku: cyklus močoviny
Ryby likvidujú amoniak tak, že ho vložia do vody pomocou žiabier ketoglutarát a amoniak, ale nepovedali sme, že k tomu dochádza iba v mitochondriách pečene.
Mitochondriálne transaminázy hrajú zásadnú úlohu v odstraňovaní amoniaku prostredníctvom cyklu močoviny.

Oxid uhličitý vo forme hydrogénuhličitanového iónu (HCO3-) sa aktivuje biotínovým kofaktorom za vzniku karboxy biotínu, ktorý reaguje s amoniakom za vzniku kyseliny karbamovej; v ďalšej reakcii sa používa ATP na prenos fosfátu na karbamylfosfát a karbamylfosfát za vzniku kyseliny karbamovej (premena ATP na ADP je hybnou silou získavania karboxybiotínu). Táto fáza je katalyzovaná karbamylfosfátsyntetáza a vyskytuje sa v mitochondriách. Karbamylfosfát a ornitín sú substráty pre enzým ornitín trans karbamyláza ktorý ich prevádza na citrulín; k tejto reakcii dochádza v mitochondriách (hepatocytoch). Produkovaný citrulín opúšťa mitochondrie a v cytoplazme podlieha „pôsobeniu“arginínsukcinátsyntetáza: dochádza k fúzii medzi uhlíkatou kostrou citrulínu a aspartátu prostredníctvom nukleofilného útoku a následnej eliminácie vody. Enzým arginínsukcinátsyntetáza vyžaduje molekulu ATP, takže dochádza k energetickému spojeniu: hydrolýza ATP na AMP a pyrofosfát (ten sa potom premení na dve molekuly ortofosfátu) nastáva vytlačením molekuly d “vody zo substrátu a nie pôsobením vody média.
„Ďalší enzým je“arginínsukcináza: tento enzým je schopný rozdeliť arginínsukcinát na arginín a fumarát v cytoplazme.
Cyklus močoviny je ukončený enzýmom argináza: získa sa močovina a ornitín; močovina sa likviduje obličkami (močom), zatiaľ čo ornitín sa vracia do mitochondrií a pokračuje v cykle.
Cyklus močoviny podlieha nepriamej modulácii arginínom: akumulácia arginínu naznačuje, že cyklus močoviny sa musí urýchliť; modulácia arginínu je nepriama, pretože arginín pozitívne moduluje enzým acetyl glutamát syntetázy. Ten je schopný prenášať acetylovú skupinu na dusík glutamátu: Vytvorí sa N-acetyl glutamát, ktorý je priamym modulátorom enzýmu karbamyl-fosfo syntetázy.
Arginín sa hromadí ako metabolit močovinového cyklu, ak produkcia karbamylfosfátu nestačí na zneškodnenie ornitínu.
Močovina sa produkuje iba v pečeni, ale existujú aj iné miesta, kde prebiehajú počiatočné reakcie.
Mozog a svaly používajú špeciálne stratégie na elimináciu aminoskupín. Mozog používa veľmi efektívnu metódu, pri ktorej sa používa enzým glutamín syntetáza a enzým glutamáza: prvý je prítomný v neurónoch, zatiaľ čo druhý sa nachádza v pečeni. Tento mechanizmus je veľmi účinný z dvoch dôvodov:
Dve aminoskupiny sú transportované z mozgu do pečene v jednom vehikule;
Glutamín je oveľa menej toxický ako glutamát (glutamát tiež vykonáva prenos neurónov a nesmie prekročiť fyziologickú koncentráciu).
V rybách podobný mechanizmus prináša aminoskupinu aminokyselín do žiabrov.
Zo svalu (kostrového a srdcového) sa aminoskupiny dostanú do pečene prostredníctvom cyklu glukóza-alanín; zapojeným enzýmom je glutamin-pyruvát transamináza: umožňuje transpozíciu aminoskupín (ktoré sú vo forme glutamátu), premenu pyruvátu na alanín a súčasne glutamátu na α-ketoglutarát vo svale a katalyzáciu reverzný proces v pečeni.
Transaminázy s rôznymi úlohami alebo polohami majú tiež štrukturálne rozdiely a dajú sa určiť elektroforézou (majú rôzne izoelektrické body).
Prítomnosť transamináz v krvi môže byť symptómom poškodenia pečene alebo srdca (tj. Poškodenia tkaniva pečeňových alebo srdcových buniek); transaminázy sú vo veľmi vysokých koncentráciách tak v pečeni, ako aj v srdci: elektroforézou je možné zistiť, či k poškodeniu došlo v pečeňových alebo srdcových bunkách.

.jpg)
.jpg)